G - پروتئین ها سیگنال ارسالی را تقویت می کنند. به عنوان مثال، نوراپی نفرین فرستنده تکانه عصبی می تواند تنها در چند میلی ثانیه با گیرنده غشایی خود تعامل کند. G - پروتئین مدت زمان سیگنال را از میلی ثانیه به ده ها ثانیه افزایش می دهد که بسیار مهم است (نیازی به ارسال مداوم سیگنال به سیستم عصبی نیست). صرفه جویی در انرژی عصبی وجود دارد.
گیرنده های جفت شده با پروتئین G خانواده گیرنده های "سرپانتین" (یا مار) را تشکیل می دهند که به این دلیل نامیده می شوند زیرا زنجیره های پلی پپتیدی آنها 7 بار از غشای پلاسمایی عبور می کنند.
این خانواده شامل گیرنده های آمین های آدرنرژیک، سروتونین، استیل کولین (موسکارینی)، بسیاری از هورمون های پپتیدی، اپیتلیوم بویایی، گیرنده های بینایی (در مخروط ها و میله های شبکیه) می باشد. مولکول اطلاعات (به عنوان مثال، نوراپی نفرین) به "جیب" تشکیل شده توسط مناطق گذرنده گیرنده متصل می شود. تغییرات حاصل در ترکیب این نواحی به حلقه های سیتوپلاسمی گیرنده منتقل می شود که پروتئین G را فعال می کند. هر چه مولکول های آگونیست بیشتر باشد، سرعت اتصال آن به گیرنده بیشتر می شود.
حساسیت زدایی گیرنده ها
این به این معنی است که پس از رسیدن به سطح بالای اولیه اولیه (مثلاً تجمع cAMP درون سلولی، جریان Na +، انقباض عضلانی و غیره)، پاسخ سلول به تدریج در طی یک دوره زمانی چند ثانیه یا دقیقه کاهش مییابد، حتی با وجود حضور مداوم سیگنالها. مولکول حساسیت زدایی برگشت پذیر است. بنابراین، 15 دقیقه پس از حذف مولکول سیگنال، قرار گرفتن در معرض مکرر آن منجر به واکنشی می شود که از نظر بزرگی با واکنش اولیه قابل مقایسه است.
پایین - تنظیم گیرنده ها.
گیرنده، زمانی که بیش از حد تحریک شود، می تواند در سیتوزول فرو رود و سلول، با کمک آنزیم های لیزوزومی، آن را به اسیدهای آمینه "هضم" می کند. غشایی که گیرنده در آن قرار داشت بازسازی می شود.
Ar – تنظیم.
اگر اعصاب عصب دهی کننده عضله با جراحی بریده شوند، عضله سیگنال هایی از سیستم عصبی دریافت نمی کند و نمی تواند منقبض شود. پاسخ عضلانی به عصب کشی با هدف سنتز گیرنده های اضافی است. آنها سنتز شده و در غشای سلولی بیرونی قرار می گیرند. سلول می خواهد سیگنالی برای انقباض دریافت کند. سیگنال نمی رسد (عصب قطع می شود)، اگرچه گیرنده های زیادی وجود دارد و به ویژه در برابر انتقال دهنده عصبی حساس هستند. گیرنده ها حتی در مکان های دیگر و دور از محل اتصال عصب به عضله قرار دارند. این به اصطلاح AP - تنظیم گیرنده ها - سنتز گیرنده های جدید توسط سلول و ادغام آنها در غشاء است. گیرنده ها دائما به روز می شوند. طول عمر گیرنده چندین روز است. به جای آن که پیر شده و توسط سلول ویران شده است، یک سلول جدید می سازد. این یک فرآیند پویا است.
بنابراین، گیرنده تعبیه شده در غشاء یک سیگنال (تکانه عصبی، هورمون دارویی) دریافت می کند و پروتئین G این سیگنال را تقویت می کند. عنصر مؤثر (آنزیم) این سیگنال را اجرا می کند و باعث سنتز پیام رسان های دوم در سلول می شود. آنها سرعت واکنش های بیوشیمیایی را در سلول ها تغییر می دهند و مستقیماً سیگنال ارسال شده توسط سیستم عصبی یا هورمونی را اجرا می کنند.
واسطه های ثانویه
1) اردوگاه.در انتقال اثرات هورمونی مانند: 1) بسیج ذخایر انرژی (تجزیه کربوهیدرات ها در کبد یا تری گلیسیرید در سلول های چربی - اثرات کاتکول آمین ها– (اپی نفرین، ایزوپرنالین).
2) احتباس آب توسط کلیه ها - اثرات وازوپرسین؛
3) حفظ هموستاز Ca +2 - اثرات هورمون های پاراتیروئید.
4) افزایش دفعات و قدرت انقباضات عضله قلب - اثرات کاتکول آمین ها (اپی نفرین، ایزوپرنالین)
5) تنظیم بیوسنتز استروئید در غدد آدرنال و غدد جنسی - اثرات کورتیکوتروپین یا هورمون محرک فولیکول.
6) – شل شدن عضلات صاف و بسیاری از فرآیندهای هورمونی و عصبی دیگر.
هنگامی که محرک عصبی یا هورمونی کامل شد، اثرات درون سلولی cAMP با فعال شدن آنزیم تخریب کننده cAMP خاتمه می یابد.
یکی از مکانیسم های عمل درمانی کافئین, تئوفیلینو سایر متیل گزانتین ها مانع از تجزیه cAMP می شوند.
2) Ca +2 و فسفوئینوزیتیدها.
برخی از هورمونها، انتقالدهندههای عصبی و فاکتورهای رشد به گیرندهای روی سطح سلول مؤثر متصل میشوند. سیگنال به پروتئین G منتقل می شود. متعاقباً فسفولیپاز C فعال می شود که فسفولیپیدهای غشای پلاسمایی را با تشکیل دو پیام رسان دوم به طور خاص می شکافد: 1. دی اسیل گلیسرول, 2) اینوزیتول تری فسفات.
دی اسیل گلیسرولپروتئین کیناز C را فعال می کند که آنزیم ها را فسفریله کرده و فعالیت آنها را تغییر می دهد.
اینوزیتول تری فسفات Ca 2+ را از ذخایر داخل سلولی (شبکه سارکوپلاسمی، میتوکندری) آزاد می کند. Ca 2+ عملکرد سلول را تغییر می دهد. مثلاً انقباض عضلانی و غیره را تحریک می کند).
لیتیوم که برای درمان حالت های شیدایی- افسردگی استفاده می شود، از طریق فسفوئینوزیتیدها عمل می کند.
3) cGMP. برخلاف cAMP، فقط در انواع خاصی از سلول ها در انتقال سیگنال نقش دارد. در مخاط روده و عضلات صاف عروق موازی با سیستم cAMP (به عنوان یک سیستم ذخیره) عمل می کند. مکانیسم اثر cGMP نیز با فسفوریلاسیون پروتئین انجام می شود.
افزایش غلظت cGMP باعث شل شدن عضله صاف عروق به دلیل دفسفوریلاسیون زنجیره های سبک میوزین می شود.
فسفوریلاسیون: مکانیسم عمومی
تقریباً تمام مکانیسم های انتقال سیگنال از طریق پیام رسان های دوم شامل فسفوریلاسیون هستند.
در طول فرآیند تکامل، بدن گیرنده های خاصی برای داروها ایجاد نکرد. آنها از طریق گیرنده های انتقال دهنده های عصبی و هورمون ها عمل می کنند. تقریباً همه داروها (به استثنای بیهوشی عمومی) اثر خود را از طریق گیرنده ها اعمال می کنند.
ما به طور مفصل گیرنده های ساخته شده در غشای پلاسمایی سلول را بررسی کردیم. اما گیرنده های دارویی دیگری نیز وجود دارد. اساساً یک گیرنده چیزی است که دارو در بدن با آن پیوند برقرار می کند (تعامل می کند). به عنوان مثال، آلبومین یک گیرنده برای داروهایی است که به آن متصل می شود. اما این گیرنده فعال نیست و منجر به اثر دارویی نمی شود.
سایر کلاس های گیرنده های دارویی عبارتند از:
1) آنزیم ها، 2) پروتئین های انتقال دهنده، 3) پروتئین های ساختاری.
هنگامی که به داروها متصل می شوند، می توان آنها را مهار کرد یا (به طور معمول کمتر) فعال کرد. به عنوان مثال، دی هیدروفولات ردوکتاز گیرنده ای برای متوترکسات
پروتئین های انتقال دهنده(به عنوان مثال، گیرنده غشایی برای گلیکوزیدهای قلبی - Na +، K +، ATPase).
پروتئین های ساختاری(به عنوان مثال، توبولین - یک گیرنده برای داروهای ضد التهابی کلشی سین).
در هر مورد از تعامل یک دارو با یک گیرنده، یک مجموعه دارو-گیرنده تشکیل می شود که منجر به تغییر در متابولیسم در سلول و اندام می شود. یک اثر دارویی ایجاد می شود. مقدار آن متناسب با تعداد کمپلکس های گیرنده دارو است.
داروهایی که عملکرد آنها با تحریک گیرنده ها همراه است نامیده می شوند آگونیست ها. آگونیست ها عبارتند از: 1) کامل (باعث پاسخ حداکثری) و 2) جزئی. دومی به گیرنده ها متصل شده و آنها را تحریک می کند. اما اثر فارماکولوژیک ضعیف تر از یک تنظیم کننده طبیعی است. موادی که در عملکرد آگونیست های خاص اختلال ایجاد می کنند، آنتاگونیست (مسدود کننده) نامیده می شوند.
گیرنده ها بر اساس حساسیت آنها به واسطه های طبیعی و آنتاگونیست های آنها طبقه بندی می شوند. به عنوان مثال، گیرنده های حساس به استیل کولین، کولینرژیک و گیرنده های حساس به اپی نفرین (آدرنالین) آدرنرژیک نامیده می شوند.
برای تحریک گیرنده ها و به دست آوردن اثر مربوطه، هم از خود واسطه ها (نوراپی نفرین، دوپامین و غیره) و هم از داروهایی که میل ترکیبی با گیرنده ها دارند استفاده می شود. اغلب، دومی آنالوگ های ساختاری واسطه ها هستند.
برخی از مواد گیرنده مربوطه را نه با برهمکنش مستقیم با آن، بلکه با آزاد کردن واسطهها از شکل محدود (از نظر فیزیولوژیکی غیرفعال) یا با مهار آنزیمهایی که واسطهها را از بین میبرند، تحریک میکنند.
گیرنده ها بخش کوچکی از غشای سلول خارجی را اشغال می کنند. بنابراین، مناطقی از غشاء که به استیل کولین واکنش نشان می دهند، تنها 1/6000 از کل سطح سلول را تشکیل می دهند.
2. تأثیر بر فعالیت آنزیم.عمل برخی از داروها بر اساس فعال یا مهار آنزیم ها است. به عنوان مثال، PHYSOSTIGMINE فعالیت کولین استراز را که استیل کولین را از بین می برد، مهار می کند. این باعث اثرات مشخصه تحریک سیستم عصبی پاراسمپاتیک می شود.
برخی از داروها می توانند باعث القاء، یعنی افزایش محتوای پروتئین آنزیم شوند. در عین حال فعالیت آنها افزایش می یابد. به عنوان مثال، فنوباربیتال با افزایش فعالیت UDP-glucuronyltransferase، هیپربیلی روبینمی را کاهش می دهد.
3. تأثیر فیزیکی و شیمیایی بر غشای سلولی.
برای برخی داروها، ماهیت مولکول های هدف ناشناخته است. عمل آنها با گیرنده های خاصی مرتبط نیست. به عنوان مثال، بیهوشی های عمومی با تغییر انتقال یون عمل می کنند. اثر درمانی پمادها، پودرها و پمادهای مایع ماهیتی فیزیکی دارد. آنها از نواحی آسیب دیده پوست یا غشاهای مخاطی از تحریک محافظت می کنند.
پروتئین های G (پروتئین های متصل شونده به GTP) واسطه های جهانی در انتقال سیگنال ها از گیرنده ها به آنزیم های غشای سلولی هستند که تشکیل پیام رسان های دوم سیگنال هورمونی را کاتالیز می کنند. پروتئین های G الیگومرهایی هستند که از زیر واحدهای α، β و γ تشکیل شده اند. ترکیب دایمرهای βγ در بافتهای مختلف کمی متفاوت است، اما در داخل یک سلول، همه پروتئینهای G، به طور معمول، مجموعهای از زیرواحدهای βγ یکسان دارند. بنابراین، پروتئین های G معمولاً با زیرواحدهای α متمایز می شوند. .
16 ژن کد کننده زیرواحد آلفا مختلف پروتئین های G شناسایی شده است. برخی از ژن ها به دلیل پیرایش RNA جایگزین، بیش از یک پروتئین دارند.
هر زیرواحد α در پروتئین G دارای مراکز خاصی است:
GTP یا GDP الزام آور؛
تعامل با گیرنده؛
اتصالات به زیر واحدهای βγ.
فسفوریلاسیون توسط پروتئین کیناز C.
برهمکنش با آنزیم آدنیلات سیکلاز یا فسفولیپاز C.
ساختار پروتئین های G فاقد حوزه های مارپیچ آلفا و پوشش غشایی است. پروتئین های G متعلق به گروه پروتئین های "لنگر" هستند (شکل 5-34).
برنج. 5-34. موقعیت پروتئین های G در غشاءبرای ارتباط پروتئین های G، اسیلاسیون پروتومرهای α با رادیکال های آلیفاتیک اسیدهای چرب، اسید میریستیک (M) یا اسید ایزوپرنیک، مهم است. زیرواحد γ پروتئین G دارای یک گروه ژرانیل-ژرانیل (G) است که توسط یک پیوند تیواستر به یک باقیمانده سیستئین C ترمینال متصل شده است.
تنظیم فعالیتجی -پروتئین ها
یک شکل غیرفعال از پروتئین G وجود دارد - کمپلکس αβγ-GDP و شکل فعال شده αβγ -GTP. فعال شدن پروتئین G پس از برهمکنش با کمپلکس فعال گیرنده اتفاق می افتد؛ تغییر در ترکیب پروتئین G میل ترکیبی زیرواحد α برای مولکول GDP را کاهش می دهد و آن را برای GTP افزایش می دهد. جایگزینی GDP با GTP در محل فعال پروتئین G، مکمل بودن بین زیر واحدهای α-GTP و βγ را مختل می کند. گیرنده مرتبط با مولکول سیگنال دهنده می تواند تعداد زیادی از مولکول های پروتئین G را فعال کند و در نتیجه سیگنال خارج سلولی را در این مرحله تقویت کند (شکل 5-35).

برنج. 5-35. چرخه عملکرد پروتئین G R s - گیرنده؛ G - هورمون؛ AC - آدنیلات سیکلاز.
زیرواحد α پروتئین G فعال شده (α-GTP) با پروتئین خاصی از غشای سلولی تعامل دارد و فعالیت آن را تغییر می دهد. چنین پروتئین هایی می توانند آنزیم های آدنیلات سیکلاز، فسفولیپاز C، cGMP فسفودی استراز، کانال های Na +، کانال های K + باشند.
مرحله بعدی چرخه عملکرد پروتئین G دفسفوریلاسیون GTP متصل به زیرواحد α است و آنزیمی که این واکنش را کاتالیز می کند خود زیرواحد α است.
دفسفوریلاسیون منجر به تشکیل یک کمپلکس α-GDP می شود که مکمل پروتئین غشایی خاص (مثلاً آدنیلات سیکلاز) نیست، اما میل ترکیبی بالایی برای پروتومرهای βγ دارد. پروتئین G به شکل غیر فعال خود یعنی αβγ-GDP باز می گردد. با فعال شدن بعدی گیرنده و جایگزینی مولکول GDP با GTP، چرخه دوباره تکرار می شود. بنابراین، زیرواحدهای α پروتئین های G برای حمل یک سیگنال محرک یا بازدارنده از یک گیرنده که توسط یک پیام رسان اولیه (مثلاً یک هورمون) فعال می شود، به آنزیمی که تشکیل یک پیام رسان ثانویه را کاتالیز می کند، حرکت می کنند.
برخی از اشکال پروتئین کینازها می توانند زیرواحدهای α پروتئین های G را فسفریله کنند. زیرواحد α فسفریله مکمل یک پروتئین غشایی خاص، به عنوان مثال، آدنیلات سیکلاز یا فسفولیپاز C نیست و بنابراین نمی تواند در انتقال سیگنال شرکت کند.
جی. آدنیلات سیکلاز
آنزیم آدنیلات سیکلاز، که تبدیل ATP به cAMP را کاتالیز می کند (شکل 5-36)، یک آنزیم کلیدی در سیستم انتقال سیگنال آدنیلات سیکلاز است. آدنیلات سیکلاز در همه انواع سلولی یافت می شود.

برنج. 5-36. تشکیل آدنوزین مونوفسفات حلقوی (cAMP).
این آنزیم متعلق به گروه پروتئین های جدایی ناپذیر غشای سلولی است و دارای 12 حوزه گذرنده است. قطعات خارج سلولی آدنیلات سیکلاز گلیکوزیله می شوند. حوزه های سیتوپلاسمی آدنیلات سیکلاز دارای دو مرکز کاتالیزوری هستند که مسئول تشکیل cAMP هستند، یک پیام رسان ثانویه که در تنظیم فعالیت آنزیم پروتئین کیناز A نقش دارد.
فعالیت آدنیلات سیکلاز تحت تأثیر تنظیم کننده های خارج سلولی و درون سلولی است. تنظیمکنندههای خارج سلولی (هورمونها، ایکوزانوئیدها، آمینهای بیوژنیک) تنظیم را از طریق گیرندههای خاصی انجام میدهند که با کمک زیرواحدهای α پروتئینهای G سیگنالها را به آدنیلات سیکلاز منتقل میکنند. αs - زیرواحد (تحریک کننده)، هنگام تعامل با آدنیلات سیکلاز، آنزیم را فعال می کند، زیرواحد αi (بازدارنده) آنزیم را مهار می کند. به نوبه خود، آدنیلات سیکلاز تظاهرات فعالیت GTP-فسفاتاز زیرواحد α را تحریک می کند. در نتیجه دفسفوریلاسیون GTP، زیر واحدهای s-GDP و i-GDP تشکیل می شوند که مکمل آدنیلات سیکلاز نیستند.
از 8 ایزوفرم مورد مطالعه آدنیلات سیکلاز، 4 ایزوفرم وابسته به Ca2+ هستند (فعال شده توسط Ca2+). تنظیم آدنیلات سیکلاز توسط کلسیم داخل سلولی به سلول اجازه می دهد تا فعالیت های دو پیام رسان اصلی cAMP و Ca 2+ را یکپارچه کند.
D. فسفولیپازها
فسفولیپازها آنزیم هایی از کلاس هیدرولاز هستند که کاتابولیسم گلیسروفسفولیپیدها را کاتابولیسم می کنند. فسفولیپازهای ترشحی که بخشی از شیره پانکراس هستند و فسفولیپازهای سلولی وجود دارند. فسفولیپازهای سلولی A 1، A 2، D، C در ویژگی آنها برای گروه بریده شده متفاوت است. تمام فسفولیپازها آنزیم های وابسته به کلسیم هستند (شکل 5-37).

برنج. 5-37. اثر فسفولیپازها
فسفولیپاز C- آنزیمی که پیوند فسفواستر را در گلیسروفسفولیپیدها هیدرولیز می کند. در سلول های انسانی، 10 ایزوفرم فسفولیپاز C شناسایی شده است که از نظر وزن مولکولی، محلی سازی، حالت تنظیم و ویژگی سوبسترا متفاوت است. ساختار تمام ایزوفرم های فسفولیپاز C فاقد حوزه های آبگریز است که می تواند برهمکنش آنها با غشاء را تضمین کند. با این حال، برخی از اشکال فسفولیپاز C با استفاده از یک "لنگر" آبگریز - باقیمانده آسیل اسید میریستیک یا به دلیل تعامل با سطح لایه دوتایی، متصل می شوند. فعالیت کاتالیزوری تمام ایزوفرم های فسفولیپاز C به یون های کلسیم بستگی دارد.
بیشتر فسفولیپازهای C برای فسفاتیدیلینوزیتول ها اختصاصی هستند و عملاً انواع دیگر فسفولیپیدها را هیدرولیز نمی کنند. آنزیم فعال می تواند تا 50 درصد از کل فسفاتیدیلینوزیتول ها را در غشای سلولی هیدرولیز کند. هیدرولیز فسفاتیدیلینوزیتول 4،5-بیس فسفات (PIF 2) محصولات دی اسیل گلیسرول (DAG) و اینوزیتول 1،4،5-تری فسفات (IP 3) را تولید می کند که به عنوان پیام رسان های ثانویه در انتقال سیگنال غشایی در طول مسیر اینوزیتول فسفات عمل می کنند.
E. پروتئین کینازها
تمام مولکولهای سیگنالدهنده قطبی که از طریق گیرندههای غشایی بر روی سلول هدف عمل میکنند، عملکرد بیولوژیکی خود را با فسفریله کردن پروتئینها و آنزیمهای خاصی که متابولیسم را در سلول تنظیم میکنند، انجام میدهند. فسفوریلاسیون باعث تغییر (افزایش یا کاهش) فعالیت آنها می شود. پروتئین کینازها فسفوریلاسیون پروتئین ها را در باقی مانده های اسید آمینه سرین، ترئونین و تیروزین کاتالیز می کنند. پروتئین کینازها ممکن است زیر واحد یک گیرنده غشایی مانند پروتئین تیروزین کیناز گیرنده انسولین باشند که فعالیت آن توسط یک هورمون تنظیم می شود. گروه دیگر پروتئین کینازهایی هستند که توسط پیام رسان های سیگنال هورمونی ثانویه تنظیم می شوند (cAMP، cGMP، Ca 2+، DAG)، به عنوان مثال پروتئین کیناز A، پروتئین کیناز C، پروتئین کیناز G، پروتئین کینازهای وابسته به کالمودولین و غیره.
پروتئین کیناز A
پروتئین کینازهای A (تحریک شده با cAMP) در سیستم انتقال سیگنال آدنیلات سیکلاز نقش دارند. پروتئین کیناز A شامل 4 زیر واحد R 2 C 2 - دو زیر واحد تنظیمی (R 2) و دو کاتالیزوری (C 2) است (شکل 5-41 را ببینید). کمپلکس R 2 C 2 فعالیت آنزیمی ندارد.
کمپلکس R 2 C 2 به طرق مختلف به غشاء متصل می شود. برخی از اشکال پروتئین کیناز A به باقی مانده اسید میریستیک آلیفاتیک زیر واحدهای کاتالیزوری متصل می شوند. در بسیاری از بافتها، پروتئین کیناز A با پروتئین «لنگر» AKAP s (از انگلیسی. پروتئینهای مهارکننده پروتئین کیناز وابسته به cAMP). ACAP s دارای یک مرکز اتصال برای زیر واحدهای تنظیمی پروتئین کیناز A است. با کمک پروتئین ACAPs، پروتئین کیناز A به غشاء در ناحیه ای که آنزیم هایی که تشکیل cAMP (آدنیلات سیکلاز) یا هیدرولیز آن را کاتالیز می کنند، متصل می شود. فسفودی استراز)، و همچنین پروتئین هایی که در تنظیم فعالیت آنزیم شرکت می کنند، به عنوان مثال، کانال های Ca2+ دارای ولتاژ.
زیرواحدهای تنظیمی پروتئین کیناز A دارای مکانهای خاصی برای اتصال cAMP هستند. اتصال cAMP به زیر واحدهای تنظیمی منجر به تغییر در ساختار دومی و کاهش میل ترکیبی برای زیر واحدهای کاتالیزوری C می شود، تفکیک طبق طرح زیر رخ می دهد:
cAMP 4 + R 2 C 2 -> cAMP 4 R 2 + C + C
زیر واحدهای C شکل فعال پروتئین کیناز A هستند که واکنشهای فسفوریلاسیون در سرین و ترئونین را کاتالیز میکنند. زیرواحدهای کاتالیزوری C انواع مختلف پروتئین کینازها یکسان نیستند، آنها عمدتاً در ویژگی آنها با توجه به پروتئین های سوبسترا متفاوت هستند.
پروتئین کیناز C
پروتئین کینازهای C در سیستم انتقال سیگنال اینوزیتول فسفات نقش دارند. آنزیم از دو حوزه عملکردی متفاوت تشکیل شده است - تنظیمی و کاتالیزوری. دامنه نظارتی شامل 2 ساختار است ("انگشت روی")توسط قطعاتی از زنجیره پپتیدی، غنی از سیستئین و حاوی 2 یون روی تشکیل شده است (به بخش 1 مراجعه کنید). انگشتان روی در اتصال دی اسیل گلیسرول نقش دارند. قطعه دیگری از حوزه تنظیمی میل ترکیبی بالایی برای Ca2+ دارد. افزایش غلظت کلسیم در سیتوزول باعث افزایش میل ترکیبی پروتئین کیناز C برای فسفاتیدیل سرین غشا می شود. انتقال پروتئین کیناز C به غشاء به آنزیم اجازه می دهد تا به DAG متصل شود، که میل پروتئین کیناز C را بیشتر برای یون های کلسیم افزایش می دهد (شکل 5-38). رایج ترین ایزوفرم های پروتئین کیناز C توسط Ca2+، دی اسیل گلیسرول و فسفاتیدیل سرین فعال می شوند.

5-38. تنظیم فعالیت پروتئین کیناز C (PKC). PS - فسفاتیدیل سرین؛ DAG - دی اسی گلیسرول.
دامنه کاتالیزوری دارای مرکزی است که ATP و پروتئین سوبسترا را متصل می کند. آنزیم فعال پروتئین کیناز C در بقایای سرین و ترئونین فسفریلات می شود. کاهش غلظت یون های کلسیم در سلول باعث اختلال در اتصال پروتئین کیناز C با فسفاتیدیل سرین و دی اسیل گلیسرول می شود، آنزیم غیرفعال می شود و از غشاء جدا می شود.
3.
پروتئین کینازها
جی
برخلاف پروتئین کیناز A، پروتئین کیناز G در همه بافت ها وجود ندارد، این پروتئین در ریه ها، مخچه، عضلات صاف و پلاکت ها یافت می شود. ایزوفرم های پروتئین کیناز G می توانند به غشاء متصل شوند یا در سیتوپلاسم قرار گیرند. پروتئین کیناز محلول C از دو زیر واحد یکسان تشکیل شده است که هر کدام دارای دو مکان برای اتصال cGMP هستند. اتصال cGMP به مراکز تنظیمی باعث تغییرات ساختاری در زیر واحدها و افزایش فعالیت کاتالیزوری آنزیم می شود (شکل 5-39). پروتئین کیناز G، مانند پروتئین کینازهای A و C، ویژه سوبستراهای پروتئینی خاصی است که در باقیمانده های سرین و ترئونین فسفریله می شود.

برنج. 5-39. تنظیم فعالیت پروتئین کیناز G (PKG).
و فسفودی استراز
فسفودی استرازها آنزیم هایی هستند که تبدیل cAMP (شکل 5-40) یا cGMP را به متابولیت های غیرفعال AMP یا GMP کاتالیز می کنند. فسفودی استرازها، با کاهش غلظت پیام رسان های ثانویه، زنجیره دگرگونی های ناشی از فعال کننده گیرنده را می شکنند.

شکل 5-40. تبدیل cAMP به AMP.
فسفودی استرازها در سلول های بافتی به دو شکل وجود دارند: پروتئین محلول و متصل به غشاء. اشکال متصل به غشاء آنزیم 5-40٪ در بافت های مختلف را تشکیل می دهند. در همان بافت، اشکال مختلفی از فسفودی استراز ممکن است وجود داشته باشد که از نظر میل ترکیبی به سوبستراها، وزن مولکولی، بار، خواص تنظیمی و محلی سازی در سلول متفاوت است.
فسفودی استرازهای نوکلئوتیدی حلقوی دارای ویژگی مطلق نیستند، بنابراین، به عنوان یک قاعده، همان شکل آنزیم قادر به هیدرولیز cAMP و cGMP است. با این حال، سرعت هیدرولیز این دو نوکلئوتید توسط فسفودی استراز یکسان می تواند به طور قابل توجهی متفاوت باشد. این بستگی به این دارد که کدام فسفودی استراز در سلول وجود دارد - مخصوص cAMP یا اختصاصی تر برای cGMP، به نسبت غلظت cAMP و cGMP در سلول و به عملکرد تنظیم کننده های فسفودی استراز.
بیشتر بافت ها حاوی فسفودی استراز-1 هستند که مخصوص cAMP است که توسط Ca2+، کمپلکس 4 Ca2+ -calmodulin و cGMP فعال می شود.
محتوا
- مقدمه 2
- از تاریخچه کشف پروتئین C 8
- ساختار و خواص 8
- اتصال غشایی 9
- سازماندهی ساختاری و عملکردی پروتئین های G 9
- طبقه بندی بر اساس حساسیت به توکن ها 10
- رابط با سیستم های افکتور 10
- تنظیم فعالیت پروتئین G 11
- آدنیلات سیکلاز 12
- فسفولیپاز 13
- پروتئین کیناز 14
- فسفودی استراز 16
- سیستم آدنیلات سیکلاز 17
- اثر سموم باکتریایی بر فعالیت آدنیلات سیکلاز (ADP-ریبوزیلاسیون پروتئین های G) 20
- سیستم فسفات اینوزیتول 21
- دخالت پروتئین کالمودولین در انتقال سیگنال اینوزیتول فسفات 22
- خود تنظیمی سیستم 23
- b-subunit: خصوصیات عمومی 23
- زیر واحدهای c و d: خصوصیات کلی 24
- G-proteins: g-sub واحد 25
- پروتئین های متصل شونده به GTP دو خانواده اصلی از پروتئین های G و پروتئین های با وزن مولکولی کم GTP را تشکیل می دهند.
- ادبیات 30
معرفی
پروتئینهای سیگنالدهنده G واسطههای جهانی در انتقال سیگنالهای هورمونی از گیرندههای غشای سلولی به پروتئینهای مؤثر هستند که باعث پاسخ سلولی نهایی میشوند. هنگامی که یک مولکول گیرنده هفت دامنه ای که در غشای یک سلول حسی قرار دارد با تغییراتی در محیط خارجی فعال می شود، دچار تغییرات ساختاری می شود. دومی توسط پروتئین های G متصل به غشاء شناسایی می شود، که به نوبه خود مولکول های موثر در غشاء را فعال می کند. این اغلب منجر به آزاد شدن پیامرسانهای دوم به داخل سیتوزول میشود. آنها به دلیل درگیری در بسیاری از فرآیندهای فیزیولوژیکی مهم، موضوع مطالعههای فشرده بودهاند. پروتئینهای G درگیر در انتقال سیگنال، اعضای یک خانواده بزرگ پروتئینهای متصل به گوانین هستند. پروتئینهای G تنظیمکنندههای دقیقی هستند که فعالیت مولکولهای دیگر را روشن یا خاموش میکنند. تقریباً 80٪ از پیامرسانهای اولیه (هورمونها، انتقالدهندههای عصبی، تعدیلکنندههای عصبی) با گیرندههای خاصی که از طریق پروتئینهای G با عوامل مؤثر مرتبط هستند، تعامل دارند. پروتئینهای G پروتئینهایی هستند که نوکلئوتیدهای گوانوزین را به هم متصل میکنند. . پروتئین های G مرتبط با گیرنده به غشاء متصل می شوند. در حالت غیر فعال، آنها با تولید ناخالص داخلی مرتبط هستند. هنگامی که گیرنده به لیگاند متصل می شود، GDP با GTP جایگزین می شود و در نتیجه فعال می شود. این فرآیند نسبتاً آهسته است و در عرض چند ثانیه رخ می دهد. آنها از یک زیرواحد b بزرگ (حدود 45 کیلو دالتون - کیلو دالتون) و همچنین زیرواحدهای b و g کوچکتر تشکیل شده اند، زیرواحد b دارای فعالیت GTPase است، به شکل غیرفعال (خاموش) مولکول تولید ناخالص داخلی را به فعال متصل می کند. سایت. زیرواحدهای c و d به هم پیوسته هستند و تحت شرایط فیزیولوژیکی قابل تفکیک نیستند. در حالت غیرفعال، کمپلکس b به طور سست به زیرواحد b متصل می شود. زیرواحد g توسط یک زنجیره ژرانیل-ژرانیل (20 اتم کربن در زنجیره) به برگ سیتوپلاسمی غشای بیولوژیکی متصل می شود که از نظر ساختار شبیه به کلسترول است. زیرواحد b نیز توسط یک اسید چرب 14 کربنی (اسید میریستوئیک) به غشاء متصل می شود. چنین اتصالاتی تضمین می کند که کمپلکس پروتئین G در صفحه غشاء نگه داشته می شود، اما در عین حال می تواند به راحتی در این صفحه حرکت کند. به راحتی می توان تصور کرد که چگونه کل مجموعه پروتئین G با GDP متصل در صفحه غشاء تحت تأثیر نیروهای حرارتی حرکت می کند؛ دو خانواده پروتئین - پروتئین های اتصال نوکلئوتید گوانوزین هتروتریمر (پروتئین های G) و گوانوزین تری فسفاتازهای مرتبط با فاصله دور (GTPases) هنگامی که GTP به اجزای پایین دست انتقال سیگنال سطح سلولی متصل می شود و فعال می شود، می توان آن را روشن کرد. GTPaseهای کوچک در کنترل خواص اساسی سلول - قطبیت شکل و فرآیندهای تقسیم و تمایز نقش دارند. پروتئین های G معمولا سیگنال های تخصصی تری را تنظیم می کنند - تولید پیام رسان های دوم. هر دو قادر به هیدرولیز GTP و در نتیجه خاموش کردن سیگنال هستند.از آنجایی که زیرواحدهای β و g پروتئین های G به شدت حفظ شده اند، پروتئین های G معمولاً با زیر واحدهای b خود متمایز می شوند. علاوه بر موتیف اتصال به GTP، هر توالی گالفا حاوی حداقل یک محل اتصال برای کاتیونهای دو ظرفیتی، و همچنین مکانهایی برای اصلاح کووالانسی توسط سموم باکتریایی است که واکنشهای ADP-ribosyltransferase وابسته به NAD را کاتالیز میکنند. پروتئینهای G که آدنیلات سیکلاز (Gs) را تحریک میکنند یا در انتقال نوری (Gt، ترانسدوسین) نقش دارند، بهعنوان سوبسترا برای ADP-ribosylation که توسط سم وبا در یکی از باقیماندههای آرژنین کاتالیز میشود، عمل میکنند که منجر به مسدود کردن غیرفعال شدن این پروتئینها میشود. Gs، پروتئین G مهارکننده آدنیلات سیکلاز (Gi) و پروتئین G با عملکرد ناشناخته (Go) توسط سم سیاه سرفه در یک باقیمانده سیستئین واقع در انتهای C با ADP ریبوزیل می شوند. این تغییر در تعامل بین پروتئین G و گیرنده ها اختلال ایجاد می کند. توالی پروتئین G موش (Gx) تعیین شد، که مشخص شد به سم سیاه سرفه حساس نیست. بهترین مورد مطالعه پروتئین های G هستند که آدنیلات سیکلاز (به ترتیب پروتئین های Gs و پروتئین های Gi) را تحریک و مهار می کنند. گیرنده های آدرنرژیک VE، گیرنده های آدرنرژیک B2 و گیرنده های D1 با پروتئین Gs جفت می شوند و بنابراین تحریک این گیرنده ها با فعال شدن آدنیلات سیکلاز و افزایش غلظت درون سلولی cAMP - پیام رسان کلاسیک دوم (داخل سلولی) همراه است. پاسخ نهایی در سلول های مختلف متفاوت است و بستگی به این دارد که قطعات موثر (آنزیم، کانال یونی و غیره) گیرنده های b2-آدرنرژیک، گیرنده های M2-کولینرژیک و گیرنده های D2 با پروتئین Gi جفت می شوند و تحریک این گیرنده ها منجر می شود. کاهش فعالیت آدنیلات سیکلاز و غلظت داخل سلولی cAMP. تغییرات در فعالیت آنزیم ها و سایر پروتئین های درون سلولی و بر این اساس، عملکردهای سلولی برخلاف آنچه در هنگام فعال شدن پروتئین Gs مشاهده می شود، است. گیرندههای β1-آدرنرژیک (مانند گیرندههای کولینرژیک M1) ظاهراً با نوع دیگری از پروتئین G مرتبط هستند که هنوز مطالعه نشده است. این پروتئین گاهی اوقات به عنوان Gq نامیده می شود. فسفولیپاز C را فعال می کند، که تجزیه فسفولیپیدهای غشایی، به ویژه فسفاتیدیلینوزیتول-4،5-دی فسفات را به ISP و DHA کاتالیز می کند. هر دوی این مواد پیام رسان های ثانویه هستند، اتصال یک آگونیست (هورمون، انتقال دهنده عصبی و غیره) به گیرنده مربوطه منجر به برهمکنش پروتئین-پروتئین بین گیرنده و پروتئین G می شود و تجزیه HDP را تسریع می کند. در نتیجه یک کمپلکس آگونیست-گیرنده-پروتئین G با عمر کوتاه تشکیل می شود که با هیچ نوکلئوتیدی مرتبط نیست. اتصال مولکول GTP به این کمپلکس باعث کاهش میل ترکیبی گیرنده برای پروتئین G می شود که منجر به تجزیه کمپلکس و آزاد شدن گیرنده می شود. به طور بالقوه، گیرنده می تواند تعداد زیادی از مولکول های پروتئین G را فعال کند، بنابراین ضریب تقویت بالای سیگنال خارج سلولی را در این مرحله فراهم می کند. زیرواحد b فعال شده پروتئین G از زیرواحدهای r جدا می شود و با عامل مربوطه برهمکنش می کند و اثر فعال کننده یا بازدارنده بر روی آن اعمال می کند. - آنزیم ها، مانند آدنیلات سیکلاز، یا احتمالا کانال های یونی. یک آنزیم می تواند فعال یا مهار شود و یک کانال یونی می تواند باز یا بسته شود. نمونه های خاص در بخش های بعدی مورد بحث قرار خواهند گرفت. با این حال، تا زمانی که زیرواحد b که یک GTPase است، GTP را حفظ کند، تعامل با اثرگذار ادامه دارد. بنابراین، خیلی زود GTP متصل به HDF هیدرولیز می شود. هنگامی که این اتفاق می افتد، زیرواحد b دوباره ترکیب خود را تغییر می دهد و توانایی خود را برای فعال کردن افکتور از دست می دهد. پس از این، b-GDP با کمپلکس r تعامل می کند و دوباره یک کمپلکس trimeric را تشکیل می دهد، بنابراین چرخه را کامل می کند. همچنین فرض بر این است که مجموعه زیرواحدهای βg نیز میتوانند (مستقیم یا غیرمستقیم) بر آنزیمهای مؤثر تأثیر بگذارند. این آنزیمها آدنیلات سیکلاز و فسفولیپاز C هستند. پروتئینهای G نیز عملکرد کانالهای یونی K و CaI+ را تنظیم میکنند. پروتئینهای G شامل پلی پپتید Gs، تحریک آدنیلات سیکلاز و تنظیم کانال های یونی CaI+، پلی پپتید Gi، مهار کننده آدنیلات سیکلاز و تنظیم کانال های K+ در سلول های بافت مغز، Gt، ترانسدوسین، دخیل در انتقال سیگنال نور، پروتئین اختصاصی گلف مژه های بویایی و غیره. همه پروتئین های G هتروتریمرهایی هستند که شامل زیرواحدهای b، c و d به ترتیب کاهش وزن مولکولی. متعاقباً، GTP مرتبط با زیرواحد b پروتئین G تحت هیدرولیز قرار می گیرد و آنزیمی که این فرآیند را کاتالیز می کند، خود زیرواحد b است. این منجر به جدا شدن زیرواحد b از عامل و ارتباط مجدد کمپلکس b-GDP با زیرواحدهای β می شود. فعال شدن خود به خودی پروتئین G مرتبط با تولید ناخالص داخلی یک فرآیند بسیار بعید است.همان مکانیسم زیربنای تنظیم هورمونی فسفولیپاز C و فسفولیپاز A2 اختصاصی فسفوئینوزیتید است. علاوه بر این، نشان داده شده است که پروتئین های G می توانند مستقیماً کانال های یونی را فعال کنند.مرحله محدود کننده سرعت در فرآیند بازگرداندن حالت اولیه پروتئین G، نرخ تفکیک GDP از زیرواحد β پروتئین G است. سرعت تفکیک زمانی که G-protein-GDP با گیرنده متصل به آگونیست تعامل می کند افزایش می یابد. اتصال GTP توسط پروتئین G ظاهرا منجر به تشکیل مجتمع پروتئین آگونیست-گیرنده-G می شود. آنالوگ GTP-STP-g-S و Mg2+ تفکیک زیرواحد b از تریمر پروتئین G را افزایش می دهد. لازم به ذکر است که زیرواحد کاتالیزوری آدنیلات سیکلاز از غشای مغز گاو از نظر کروماتوگرافی با زیرواحدهای b و b پروتئین Gs خالص سازی می شود و موضوع تفکیک زیرواحدهای b از تریمر پروتئین G برای فعال سازی عامل نیازمند است. پروتئین های G پلی مورفیسم قابل توجهی را نشان می دهند. هر شکل از زیر واحدهای پروتئین G از نظر ساختار بسیار همولوگ، از نظر عملکرد مشابه است، اما در وزن مولکولی و تحرک الکتروفورتیک متفاوت است. پلی مورفیسم به ویژه گسترده است و بیشتر برای پروتئین های bs و bi G مورد مطالعه قرار گرفته است. بنابراین، 11 شکل از DNA مسئول سنتز زیرواحدهای bs از مغز انسان جدا شده است که چهار نوع از آنها شبیه سازی شده اند و فرض بر این است که سنتز چهار ایزوفرم bs در مغز انسان تعیین می شود. برای bi، عمدتاً سه ایزوفرم یافت شد: bi1، bi2، bi3. جرم مولکولی ایزوفرم bs در محدوده 42-55 کیلو دالتون و bi 39-41 کیلو دالتون است. توزیع انواع مولکولی bi به بافت خاص است: bi1 عمدتاً در مغز ارائه می شود، bi2 در بافت عصبی و سلول های خونی یافت می شود، bi3 در بافت های محیطی وجود دارد و در مغز وجود ندارد. توزیع ژن های کد کننده سنتز سه ایزوفرم بی در بافت ها تقریباً به ترتیب زیر منطبق است: انسان، گاو، موش صحرایی، موش. تعیین توالی اسید آمینه bi و bs نشان داد که ایزوفرمهای bs یا bi در ناحیه توالیهای پایانی C و N که به گیرنده یا عامل متصل میشوند، متفاوت هستند. فرض بر این است که پلیمورفیسم زیرواحدهای b با تنوع گیرندهها و زیرشاخههای آنها و تنوع سیستمهای موثر تعیین میشود.زیر واحدهای Gi b توسط سه ژن ساختاری مختلف کدگذاری میشوند. در مورد ایزوفرمهای زیرواحد b پروتئینهای Gs، هنوز مشخص نیست که آیا ایزوفرمها توسط ژنهای ساختاری مختلف کدگذاری میشوند یا اینکه آیا آنها محصول یک ژن واحد هستند و به دنبال آن پیرایش جایگزین داخلی رونوشت RNA اصلی است یا خیر. آیا تعدد آنها نتیجه اصلاح پس از ترجمه است؟ در حال حاضر 9 ژن ساختاری کد کننده پروتئین های G و 12 محصول از این ژن ها شناخته شده است.
از تاریخچه کشف پروتئین C
1. 1971 - نیاز به GTP برای تحریک آدنیلات سیکلاز توسط گلوکاگون برای اولین بار نشان داده شد. 1981 - پروتئین Gt-transducin که رودوپسین را با فسفودی استراز با GTP گیرنده های نوری متصل می کند، جدا شده است. 1983 - Gs پروتئین متصل شونده به GTP جدا شد که گیرنده های تحریک کننده را با آدنیلات سیکلاز جفت می کند. 1985-1988 - نشان داده شد که فسفولیپاز C و فسفولیپاز A2 توسط هورمون ها و انتقال دهنده های عصبی از طریق پروتئین های Gp تنظیم می شوند. در حال حاضر، پروتئین های G به چند نوع تقسیم می شوند: چهار Gs، سه Gi، Go، Gz/x (سیستم عصبی مرکزی و طحال)، Gt (ترانسدوسین)، گلف (سلول های عصبی اپیتلیال بویایی).
ساختار و خواص
1. پروتئین های G هتروتریمرهایی هستند که در آنها زیرواحد b به طور سست به دایمر c-g متصل است.2. همه زیرواحدهای b شناخته شده (مول. جرم - 50 کیلو دالتون) همولوگ هستند و اکثر آنها زیرواحدهای b یکسان (یا بسیار مشابه) (مول. جرم 35 کیلو دالتون) و زیرواحدهای g (مول. جرم 8 کیلو دالتون) دارند. زیرواحد b ویژگی اتصال پروتئین G به گیرنده و عامل را تعیین می کند و برای هر پروتئین G منحصر به فرد است. زیرواحد b GTP (GTPase) را متصل و هیدرولیز می کند.5. زیرواحد b حاوی یک دامنه اتصال و هیدرولیز GTP بسیار حفاظت شده است (18 اسید آمینه از 350-395).6. مکانهای اتصال برای نوکلئوتیدهای گوانین و مکانهای برهمکنش با گیرندهها (C-پایانه) و g-dimers (N-پایانه) شناسایی شدهاند.7. محل های ADP-ریبوزیلاسیون (آرژنین-202) تحت تأثیر سم وبا و سم سیاه سرفه شناسایی شده است.
اتصال غشایی
پروتئین های G در سطح داخلی غشای پلاسمایی قرار دارند. ساختار اولیه همه زیرواحدهای پروتئین G شامل حوزههای آبگریز و پوشاننده غشا نیست. ارتباط پروتئین های G با غشاء با اسیلاسیون توسط رادیکال های اسید چرب تسهیل می شود. دو نوع تغییر لیپیدی زیرواحدهای پروتئین G شناسایی شده است: myristoylation و isoprenylation زنجیره پروتئین. myristoylation پس از ترجمه در انتهای N برای زیرواحدهای b پروتئین Go و Gi نشان داده شده است. تغییرات پس از ترجمه (acylation) نیز برای زیر واحدهای rg نشان داده شده است. سه تغییر متوالی پس از ترجمه مسئول اتصال پروتئینهای ras به غشاء شناسایی شدهاند. زیرواحدهای β خالص شده خاصیت آبدوستی از خود نشان می دهند (بدون کمپلکس β نمی توانند به وزیکول های فسفولیپید مصنوعی متصل شوند).
سازماندهی ساختاری و عملکردی پروتئین های G
پروتئین های G (پروتئین های متصل شونده به GTP) واسطه های جهانی در انتقال سیگنال ها از گیرنده ها به آنزیم های غشای سلولی هستند که تشکیل پیام رسان های دوم سیگنال هورمونی را کاتالیز می کنند. پروتئین های G الیگومرهایی هستند که از زیر واحدهای b، c و g تشکیل شده اند. ترکیب دایمرهای Gg در بافت های مختلف کمی متفاوت است، اما در داخل یک سلول، همه پروتئین های G، به طور معمول، دارای مجموعه ای از زیر واحدهای Gg هستند. بنابراین، پروتئین های G معمولاً با زیرواحدهای b خود متمایز می شوند. 16 ژن کد کننده زیر واحدهای مختلف b از پروتئین های G شناسایی شده است. برخی از ژن ها به دلیل پیرایش RNA جایگزین، بیش از یک پروتئین دارند. هر زیرواحد α در پروتئین G مراکز خاصی دارد: GTP یا GDP اتصال، برهمکنش با گیرنده، اتصال به زیرواحدهای β، فسفوریلاسیون تحت اثر پروتئین. کیناز C؛ برهمکنش با آنزیم آدنیلات سیکلاز یا فسفولیپاز C. ساختار پروتئینهای G فاقد حوزههای b-مارپیچ و پوشش غشایی است. پروتئین های G متعلق به گروه پروتئین های "لنگر" هستند.
طبقه بندی بر اساس حساسیت به توکن ها
1. CT (سم وبا) منجر به فعال شدن مداوم آدنیلات سیکلاز (سرکوب فعالیت GTPase زیر واحد As) می شود. CT (سم سیاه سرفه) همچنین باعث ریبوزیلاسیون ADP زیر واحد b می شود. علاوه بر این، در این مورد، اصلاح پروتئین G از تعامل آن با گیرنده ها جلوگیری می کند، بنابراین، زمانی که گیرنده A فعال می شود، مهار نمی شود. با توجه به حساسیت به سموم وبا و سیاه سرفه، پروتئین های G را می توان به چهار گروه تقسیم کرد: فقط حساس. به سم وبا (Gs)، فقط به سیاه سرفه (Gi و Go)، سوبستراهای هر دو سم (Gt) و پروتئین های G، که زیرواحدهای b آنها به هیچ کدام از سموم حساس نیستند.
رابط با سیستم های اثرگذار
پروتئین های متصل شونده به GTP چندین آنزیم غشایی و تعدادی کانال یونی را کنترل می کنند.اسکلت سلولی احتمالاً با پروتئین های G برهمکنش می کند و به همین دلیل هورمون ها ترشح و اندوسیتوز را تنظیم می کنند. از اهداف غشایی و درون سلولی پروتئین های G، بهترین مورد مطالعه شده آدنیلات سیکلاز و فسفودی استراز GMP شبکیه است که به ترتیب توسط Gzx و ترانسدوسین فعال می شوند. این دو آنزیم از نظر ساختار و مکانیسم تنظیم آنها توسط پروتئین های G تفاوت اساسی با یکدیگر دارند.در مورد فعال شدن سایر سیستم های وابسته به پروتئین G هیچ شفافیتی وجود ندارد. پروتئینهای G نهتنها در اثر فعالسازی، بلکه در اثر بازدارندگی آگونیستها بر روی سیستمهای موثر درون سلولی نیز نقش دارند. مهار وابسته به G-protein برای آدنیلات سیکلاز، کانال های کلسیمی با ولتاژ، فسفولیپاز C، Na/K-ATPase نشان داده شده است. . یکی از آنها به دلیل عملکرد زیرواحدهای g است و ظاهراً برای همه پروتئین های G یکسان است، زیرا زیرواحدهای g آنها مشابه هستند. مکانیسم دوم مهار ویژه آدنیلات سیکلاز توسط زیرواحد b پروتئین Gi است.
تنظیم فعالیت پروتئین G
یک شکل غیرفعال از پروتئین G وجود دارد - کمپلکس bvg-GDP و شکل فعال شده bvg-GTP. فعال شدن پروتئین G پس از برهمکنش با کمپلکس فعال گیرنده رخ می دهد؛ تغییر در ترکیب پروتئین G میل ترکیبی زیرواحد b را برای مولکول GDP کاهش می دهد و آن را برای GTP افزایش می دهد. جایگزینی GDP با GTP در فعال مرکز پروتئین G، مکمل بودن بین زیر واحدهای b-GTP و g-G را مختل می کند. گیرنده مرتبط با مولکول سیگنال دهنده می تواند تعداد زیادی مولکول پروتئین G را فعال کند، در نتیجه سیگنال خارج سلولی را در این مرحله تقویت می کند. . چنین پروتئین هایی می توانند آنزیم های آدنیلات سیکلاز، فسفولیپاز C، cGMP فسفودی استراز، کانال های Na+، کانال های K+ باشند. مرحله بعدی چرخه عملکرد پروتئین G، دفسفوریلاسیون GTP مرتبط با زیرواحد b است و آنزیمی که این واکنش را کاتالیز می کند، خود زیرواحد b. زیرواحد دفسفوریلاسیون منجر به تشکیل کمپلکس b-GDP می شود که مکمل پروتئین غشایی خاص (مثلاً آدنیلات سیکلاز) نیست، اما میل ترکیبی بالایی برای بتا پروتومرها دارد. پروتئین G به شکل غیر فعال خود یعنی bvg-GDF باز می گردد. با فعال شدن بعدی گیرنده و جایگزینی مولکول GDP با GTP، چرخه دوباره تکرار می شود. بنابراین، زیرواحدهای β پروتئینهای G جابهجا میشوند و یک سیگنال محرک یا بازدارنده را از گیرندهای که توسط یک پیامرسان اولیه (مثلاً یک هورمون) فعال میشود، به آنزیمی که تشکیل یک پیامرسان ثانویه را کاتالیز میکند، منتقل میکنند. برخی از اشکال پروتئین. کینازها می توانند زیرواحدهای β پروتئین های G را فسفریله کنند. زیرواحد b فسفریله مکمل یک پروتئین غشایی خاص مانند آدنیلات سیکلاز یا فسفولیپاز C نیست و بنابراین نمی تواند در انتقال سیگنال شرکت کند.
آدنیلات سیکلاز
آنزیم آدنیلات سیکلاز، که تبدیل ATP به cAMP را کاتالیز می کند، یک آنزیم کلیدی در سیستم انتقال سیگنال آدنیلات سیکلاز است. آدنیلات سیکلاز در همه انواع سلول ها یافت می شود.این آنزیم از گروه پروتئین های جدایی ناپذیر غشای سلولی است که دارای 12 حوزه گذرنده است. قطعات خارج سلولی آدنیلات سیکلاز گلیکوزیله می شوند. حوزههای سیتوپلاسمی آدنیلات سیکلاز دارای دو مرکز کاتالیزوری هستند که مسئول تشکیل cAMP هستند، پیامرسان دومی که در تنظیم فعالیت آنزیم پروتئین کیناز A نقش دارد. فعالیت آدنیلات سیکلاز تحت تأثیر تنظیمکنندههای خارج سلولی و درون سلولی است. تنظیمکنندههای خارج سلولی (هورمونها، ایکوزانوئیدها، آمینهای بیوژنیک) تنظیم را از طریق گیرندههای خاصی انجام میدهند که با استفاده از زیرواحدهای b پروتئینهای G، سیگنالها را برای آدنیلات سیکلاز ارسال میکنند. زیرواحد bs (تحریک کننده)، هنگام تعامل با آدنیلات سیکلاز، آنزیم را فعال می کند، دو زیرواحد (بازدارنده) آنزیم را مهار می کند. به نوبه خود، آدنیلات سیکلاز تظاهرات فعالیت GTP فسفاتاز زیرواحدهای b را تحریک می کند. در نتیجه دفسفوریلاسیون GTP زیرواحدهای bs-GDP و bi-GDP تشکیل می شوند که غیر مکمل آدنیلات سیکلاز هستند.از 8 ایزوفرم مورد مطالعه آدنیلات سیکلاز، 4 ایزوفرم وابسته به Ca2+ (فعال شده توسط Ca2+) هستند. تنظیم آدنیلات سیکلاز توسط کلسیم داخل سلولی به سلول اجازه می دهد تا فعالیت های دو پیام رسان اصلی cAMP و Ca2+ را یکپارچه کند.
فسفولیپازها
فسفولیپازها آنزیم هایی از کلاس هیدرولاز هستند که کاتابولیسم گلیسروفسفولیپیدها را کاتابولیسم می کنند. فسفولیپازهای ترشحی که بخشی از شیره پانکراس هستند و فسفولیپازهای سلولی وجود دارند. فسفولیپازهای سلولی A1، A2، D، C در ویژگی آنها برای گروه بریده شده متفاوت است. تمام فسفولیپازها آنزیم های وابسته به کلسیم هستند فسفولیپاز C آنزیمی است که پیوند فسفواستر موجود در گلیسروفسفولیپیدها را هیدرولیز می کند. در سلول های انسانی، 10 ایزوفرم فسفولیلاز C شناسایی شده است که از نظر وزن مولکولی، محلی سازی، حالت تنظیم و ویژگی سوبسترا متفاوت است. ساختار تمام ایزوفرم های فسفولیپاز C فاقد حوزه های آبگریز است که می تواند برهمکنش آنها با غشاء را تضمین کند. در این مورد، برخی از اشکال فسفولیپاز C با استفاده از یک "لنگر" آبگریز - باقیمانده آسیل اسید میریستیک یا به دلیل تعامل با سطح لایه پشت سر هم با غشاء مرتبط می شوند. فعالیت کاتالیزوری تمام ایزوفرم های فسفولیپاز C به یون های کلسیم بستگی دارد.بیشتر فسفولیپازهای C برای فسفاتیدیلینوزیتول ها اختصاصی هستند و عملاً انواع دیگر فسفولیپیدها را هیدرولیز نمی کنند. آنزیم فعال می تواند تا 50 درصد از کل فسفاتیدیلینوزیتول ها را در غشای سلولی هیدرولیز کند. هیدرولیز فسفاتیدیلینوزیتول 4،5-بیس فسفات (PIP2) باعث تولید هیدروکربن های دی اسیل گلیسرول (DAG) و اینوزیتول 1،4،5-تری فسفات (IP3) می شود که به عنوان پیام رسان ثانویه در انتقال سیگنال غشایی در طول مسیر اینوزیتول فسفات عمل می کنند.
پروتئین کینازها
تمام مولکولهای سیگنالدهنده قطبی که از طریق گیرندههای غشایی بر روی سلول هدف عمل میکنند، عملکرد بیولوژیکی خود را با فسفریله کردن پروتئینها و آنزیمهای خاصی که متابولیسم را در سلول تنظیم میکنند، انجام میدهند. فسفوریلاسیون باعث تغییر (افزایش یا کاهش) فعالیت آنها می شود. پروتئین کینازها فسفوریلاسیون پروتئین ها را در باقی مانده های اسید آمینه سرین، ترئونین و تیروزین کاتالیز می کنند. پروتئین کینازها ممکن است زیر واحد یک گیرنده غشایی مانند پروتئین تیروزین کیناز گیرنده انسولین باشند که فعالیت آن توسط یک هورمون تنظیم می شود. گروه دیگر پروتئین کینازهایی هستند که توسط پیام رسان های سیگنال هورمونی ثانویه تنظیم می شوند (cAMP، cGMP، Ca، DAG)، برای مثال پروتئین کیناز A، پروتئین کیناز C، پروتئین کیناز G، پروتئین کینازهای وابسته به کالمودولین، و غیره. پروتئین کینازها Aپروتئین کینازهای A (تحریک شده با cAMP) در سیستم انتقال سیگنال آدنیلات سیکلاز نقش دارند. پروتئین کیناز A از 4 زیر واحد R2C2 - دو زیر واحد تنظیمی (R2) و دو کاتالیزوری (C2) تشکیل شده است. کمپلکس R2C2 فعالیت آنزیمی ندارد کمپلکس R2C2 به طرق مختلف به غشاء متصل می شود. برخی از اشکال پروتئین کیناز A به باقی مانده اسید میریستیک آلیفاتیک زیر واحدهای کاتالیزوری متصل می شوند. در بسیاری از بافتها، پروتئین کیناز A با پروتئین AKAPs "لنگردار" (از پروتئینهای مهارکننده پروتئین کیناز وابسته به cAMP انگلیسی) مرتبط است. AKAPها دارای یک مرکز اتصال برای زیر واحدهای تنظیمی پروتئین کیناز A هستند. با کمک پروتئین AKAPs، پروتئین کیناز A به غشاء در ناحیه ای که آنزیم ها تشکیل cAMP (آدنیلات سیکلاز) یا هیدرولیز آن (فسفودی استراز) را کاتالیز می کنند، متصل می شود. و همچنین پروتئین هایی که در تنظیم فعالیت آنزیم شرکت می کند، به عنوان مثال کانال های Ca2+ دارای ولتاژ. زیرواحدهای تنظیم کننده پروتئین کیناز A دارای مراکز خاصی برای اتصال cAMP هستند. اتصال cAMP به زیر واحدهای تنظیمی منجر به تغییر در ساختار دومی و کاهش میل ترکیبی برای زیرواحدهای کاتالیزوری C می شود، تفکیک طبق این طرح رخ می دهد: cAMP4 + R2C2 = cAMP4R2 + C + CC زیر واحدهای C شکل فعال هستند. پروتئین کیناز A، که واکنش های فسفوریلاسیون پروتئین ها در سرین و ترئونین را کاتالیز می کند. زیرواحدهای کاتالیزوری C انواع مختلف پروتئین کینازهای A یکسان نیستند؛ آنها عمدتاً در ویژگی آنها با توجه به پروتئین های سوبسترا متفاوت هستند. پروتئین کینازهای C پروتئین کینازهای C در سیستم انتقال سیگنال اینوزیتول فسفات نقش دارند. آنزیم از دو حوزه عملکردی متفاوت تشکیل شده است - تنظیمی و کاتالیزوری. دامنه تنظیمی شامل 2 ساختار ("انگشت روی") است که توسط قطعات زنجیره پپتیدی غنی از سیستئین و حاوی دو یون روی تشکیل شده است. "انگشتان روی" در اتصال: دی اسیل گلیسرول نقش دارد. قطعه دیگری از حوزه تنظیمی میل ترکیبی بالایی برای Ca2 + دارد. افزایش غلظت کلسیم در سیتوزول باعث افزایش میل ترکیبی پروتئین کیناز C برای فسفاتیدیل سرین غشا می شود. انتقال پروتئین کیناز C به غشاء به آنزیم اجازه می دهد تا به DAG متصل شود، که میل پروتئین کیناز C را بیشتر برای یون های کلسیم افزایش می دهد. رایج ترین ایزوفرم های پروتئین کیناز C توسط Ca2+، دی اسیل گلیسرول و فسفاتیدیل سرین فعال می شوند.دامنه کاتالیزوری دارای مرکزی است که ATP و پروتئین سوبسترا را متصل می کند. شکل فعال آنزیم پروتئین کیناز C پروتئین ها را در باقی مانده های سرین و ترئونین فسفریله می کند. کاهش غلظت یون های کلسیم در سلول باعث اختلال در اتصال پروتئین کیناز C با فسفاتیدیل سرین و دی اسیل گلیسرول می شود، آنزیم غیرفعال شده و از غشاء جدا می شود.3. پروتئین کیناز G برخلاف پروتئین کیناز A، پروتئین کیناز G در همه بافت ها وجود ندارد، این پروتئین در ریه ها، مخچه، عضلات صاف و پلاکت ها یافت می شود. ایزوفرم های پروتئین کیناز G می توانند به غشاء متصل شوند یا در سیتوپلاسم قرار گیرند. پروتئین کیناز G محلول از دو زیر واحد یکسان تشکیل شده است که هر کدام دارای دو مکان برای اتصال cGMP هستند. اتصال cGMP به مراکز تنظیمی باعث تغییرات ساختاری در زیر واحدها و افزایش فعالیت کاتالیزوری آنزیم می شود. پروتئین کیناز G، مانند پروتئین کینازهای A و C، ویژه سوبستراهای پروتئینی خاصی است که در باقیمانده های سرین و ترئونین فسفریله می شود.
فسفودی استرازها
فسفودی استرازها آنزیم هایی هستند که تبدیل cAMP یا cGMP را به متابولیت های غیر فعال AMP یا GMP کاتالیز می کنند. فسفودی استرازها با کاهش غلظت پیام رسان های ثانویه، زنجیره دگرگونی های ناشی از فعال گیرنده را می شکنند. اشکال متصل به غشاء آنزیم 5-40٪ در بافت های مختلف را تشکیل می دهند. در یک بافت، اشکال مختلف فسفودی استراز ممکن است وجود داشته باشد، که از نظر میل ترکیبی به سوبستراها، وزن مولکولی، بار، خواص تنظیمی و محلی سازی در سلول متفاوت است. این آنزیم قادر به هیدرولیز cAMP و cGMP است. علاوه بر این، سرعت هیدرولیز این دو نوکلئوتید تحت اثر فسفودی استراز یکسان می تواند به طور قابل توجهی متفاوت باشد. این بستگی به این دارد که کدام فسفودی استراز در سلول وجود دارد - مخصوص cAMP یا اختصاصی تر برای cGMP، به نسبت غلظت cAMP و cGMP در سلول و به عملکرد تنظیم کننده های فسفودی استراز. در اکثر بافت ها، فسفودی استراز 1 وجود دارد. اختصاصی تر به cAMP، Ca2+ فعال شده، کمپلکس 4Ca2+-calmodulin و cGMP.
سیستم آدنیلات سیکلاز
با مشارکت سیستم آدنیلات سیکلاز، اثرات صدها مولکول سیگنالینگ مختلف هورمون ها، انتقال دهنده های عصبی و ایکوزانوئیدها مشخص می شود. عملکرد سیستم انتقال سیگنال از طریق غشاء توسط پروتئین ها تضمین می شود: گیرنده Rs مولکول سیگنال، که آدنیلات سیکلاز و گیرنده ری مولکول سیگنال را فعال می کند که آدنیلات سیکلاز را مهار می کند. پروتئین های محرک Gs و Gi-adenylate سیکلاز. آنزیم های آدنیلات سیکلاز (AC) و پروتئین کیناز A (PKA) توالی رویدادهایی که منجر به فعال شدن آدنیلات سیکلاز می شود: اتصال یک فعال کننده سیستم آدنیلات سیکلاز، به عنوان مثال، هورمون (G) به گیرنده Rs، منجر به تغییر در ساختار گیرنده و افزایش میل ترکیبی آن برای پروتئین Gs. در نتیجه، کمپلکس [G][R] تشکیل می شود؛ افزودن [G][R] به G-GDP میل ترکیبی زیرواحد b پروتئین Gs را برای تولید ناخالص داخلی کاهش می دهد و میل ترکیبی برای GTP را افزایش می دهد. تولید ناخالص داخلی با GTP جایگزین می شود؛ این امر باعث تجزیه مجموعه می شود. زیرواحد جدا شده b، مرتبط با یک مولکول GTP، میل ترکیبی با آدنیلات سیکلاز دارد: [G] [R] = [G] [R] + b-GTP + rgb برهمکنش زیرواحد b با آدنیلات سیکلاز منجر به تغییر می شود. در ترکیب آنزیم و فعالسازی آن، سرعت تشکیل cAMP از ATP را افزایش میدهد؛ تغییرات ساختاری در کمپلکس [b-GGP] [AC] باعث افزایش فعالیت GTP-فسفاتاز زیرواحد b میشود. واکنش دفسفوریلاسیون GTP رخ می دهد و یکی از محصولات واکنش، فسفات معدنی (P)، از زیر واحد b جدا می شود و کمپلکس [b-GDP] حفظ می شود. سرعت هیدرولیز هدایت سیگنال را تعیین می کند؛ تشکیل یک مولکول GDP در مرکز فعال زیرواحد b میل آن را برای آدنیلات سیکلاز کاهش می دهد، اما میل آن را برای زیرواحدهای g افزایش می دهد. پروتئین Gs به شکل غیرفعال خود باز می گردد؛ اگر گیرنده با یک فعال کننده مانند یک هورمون همراه باشد، چرخه عملکرد پروتئین Gs تکرار می شود. فعال سازی مولکول های cAMP پروتئین کیناز A (PKA) می تواند به طور برگشت پذیر به زیر واحدهای تنظیم کننده PKA متصل شود.افزودن cAMP به زیر واحدهای تنظیمی (R) باعث تجزیه کمپلکس C2R2 به کمپلکس cAMP4R2 و C+ می شود.پروتئین کیناز فعال A پروتئینهای خاص را در سرین و ترئونین فسفریله میکند و در نتیجه ساختار و فعالیت پروتئینهای فسفریله شده تغییر میکند که منجر به تغییر در سرعت و جهت فرآیندهای تنظیمکننده آنها در سلول میشود. غلظت cAMP در سلول میتواند تنظیم شده؛ بستگی به نسبت فعالیت آنزیم های آدنیلات سیکلاز و فسفودی استراز دارد.پروتئین AKAP نقش عمده ای در تنظیم سیستم سیگنال دهی داخل سلولی دارد. پروتئین "لنگر" AKAPs در جمع آوری کمپلکس های آنزیمی، از جمله نه تنها پروتئین کیناز A، بلکه فسفودی استراز و فسفوپروتئین فسفاتاز نیز نقش دارد. مکانیسم آبشاری تقویت و سرکوب سیگنال. انتقال سیگنال از گیرنده غشایی از طریق پروتئین G به آنزیم آدنیلات سیکلاز نمونه ای از سیستم آبشاری برای تقویت این سیگنال است. یک مولکول فعال کننده گیرنده می تواند چندین پروتئین G را فعال کند و هر کدام چندین مولکول آدنیلات سیکلاز را برای تولید هزاران مولکول cAMP فعال می کند. در این مرحله سیگنال 10I-10I بار تقویت می شود. cAMP حاصل توسط آنزیم دیگری به نام پروتئین کیناز A "روشن" می شود و سیگنال را 1000 بار دیگر تقویت می کند. فسفوریلاسیون آنزیم ها توسط پروتئین کیناز. و سیگنال را حتی بیشتر تقویت می کند و در نتیجه 106-107 برابر افزایش می یابد. بنابراین، با توجه به مکانیسم تقویت آبشاری، یک مولکول تنظیمکننده قادر است فعالیت میلیونها مولکول دیگر را تغییر دهد، اما برای هر یک از سیستمهای انتقال سیگنال غشایی، سلول سیستم دیگری دارد که این سیگنال را سرکوب میکند. هر یک از مراحل در آبشار آنزیمی تحت کنترل مکانیسم های خاصی است که این سیگنال را سرکوب می کند. به عنوان مثال، عملکرد طولانی مدت هورمون منجر به حساسیت زدایی گیرنده های غشایی می شود: آنها یا غیرفعال می شوند یا همراه با هورمون از طریق اندوسیتوز در سلول غوطه ور می شوند. در نتیجه حساسیت زدایی گیرنده، درجه فعال شدن سیستم آدنیلات سیکلاز کاهش می یابد. اگر غلظت cAMP در یک سلول برای مدت طولانی افزایش یابد (افزایش فعالیت پروتئین کیناز A)، فسفوریلاسیون کانال های کلسیمی ممکن است رخ دهد که منجر به افزایش غلظت CaI+ در سلول می شود. کلسیم فسفودی استراز وابسته به CaI را فعال می کند که تبدیل cAMP به AMP را کاتالیز می کند. در نتیجه غیر فعال شدن پروتئین کیناز A (R2C2)، سرعت فسفوریلاسیون آنزیم های خاص کاهش می یابد. "خاموش" سیستم توسط فسفوپروتئین فسفاتاز تکمیل می شود که فسفوپروتئین ها را دفسفریله می کند.
اثر سموم باکتریایی بر فعالیت آدنیلات سیکلاز (ADP-ریبوزیلاسیون پروتئین های G)
برای بررسی عملکرد پروتئینهای G سیستم آدنیلات سیکلاز، از سموم باکتریایی اگزوژن، سموم وبا و سیاه سرفه استفاده شد. در شرایط تجربی، سموم فعالیت آدنیلات سیکلاز را تقریباً در تمام سلولهای بدن افزایش میدهند. بنابراین، سم وبا می تواند ترشح هورمون های تیروئید توسط سلول های تیروئید، هورمون های استروئیدی توسط سلول های آدرنال و تجزیه چربی ها در سلول های چربی را تحریک کند. پاسخ سلول های مختلف به سم وبا ناشی از افزایش سطح cAMP در این سلول ها است.توکسین وبا یک پروتئین الیگومری است. یکی از زیر واحدها آنزیم ADP-ribosyltransferase است. با نفوذ به سلول، افزودن ADP-ریبوز به زیرواحد bs کمپلکس [bs - GTP] [AC] (مرحله فعالسازی آدنیلات سیکلاز) را کاتالیز میکند.NAD + + [bs, - GTP] [AC] - [ADP - ریبوسیل - bs GTP] [AC] + نیکوتین آمید + H + ADP-ریبوزیلاسیون از تظاهرات فعالیت GTP-فسفاتاز زیرواحد bs جلوگیری می کند، دفسفوریلاسیون GTP رخ نمی دهد. چرخه عملکرد پروتئین G در مرحله فعال شدن آنزیم آدنیلات سیکلاز، که مسئول تشکیل cAMP از ATP است، متوقف می شود. آنزیم آدنیلات سیکلاز فعالیت افزایش یافته را برای مدت طولانی حفظ می کند. زیرواحد سم سیاه سرفه با نفوذ به داخل سلول، ADP-ریبوزیلاسیون دو زیرواحد Gi-پروتئین فعال شده (big-GTP) را کاتالیز می کند.NAD + [big g -GTP] - [ADP-ribosyl-bi bg-GTP] + نیکوتین آمید + H + دو زیرواحد اصلاح شده میل ترکیبی بالایی را برای زیرواحدهای bg حفظ می کند. پروتئین Gi توانایی تفکیک به زیر واحدهای bi-GTP و rg را از دست می دهد. بنابراین، سیگنال بازدارنده (bi-GTP) به آدنیلات سیکلاز نمی رسد، به این معنی که در این مورد فقط فعال شدن آن با اتصال به bs-GTP امکان پذیر است. اثر سم سیاه سرفه بر سلول های بافتی همیشه منجر به افزایش سطح cAMP می شود.علائم وبا و سیاه سرفه در نتیجه عمل سموم تولید شده توسط میکروارگانیسم های مربوطه ایجاد می شود.
سیستم فسفات اینوزیتول
عملکرد سیستم انتقال سیگنال غشایی اینوزیتول فسفات توسط: R (گیرنده)، فسفولیپاز C، Gplc تضمین می شود - پروتئینی که فسفولیپاز C، پروتئین ها و آنزیم های غشاء و سیتوزول را فعال می کند. توالی وقایع منجر به فعال شدن فسفولیپاز می شود. ج: اتصال یک مولکول سیگنال دهنده، به عنوان مثال، یک هورمون به گیرنده (R ) باعث تغییر در ترکیب و افزایش میل ترکیبی برای پروتئین Gplc می شود. تشکیل کمپلکس [G][R] منجر به کاهش در میل ترکیبی b-پروتومر پروتئین Gplc برای تولید ناخالص داخلی و افزایش میل ترکیبی برای GTP. GDP جایگزین GDP می شود. زیرواحد β جدا شده، مرتبط با یک مولکول GTP، میل ترکیبی با فسفولیپاز C به دست می آورد. β-GTP با فسفولیپاز C تعامل می کند و آن را فعال می کند. تحت تأثیر فسفولیپاز C، لیپید غشایی فسفاتیدیل-4،5-بیوفسفات (PIF2) هیدرولیز می شود و در جریان هیدرولیز، ماده آبدوست اینوزیتول-1،4،5-تری فسفات (IP3) تشکیل شده و در سیتوزول آزاد می شود. محصول دیگر واکنش، دی آسیل گلیسرول (DAG) در غشاء باقی می ماند و در فعال سازی آنزیم پروتئین کیناز C (PKC) شرکت می کند.اینوزیتول-1،4،5-تری فسفات (IP3) به مراکز خاصی از کانال Ca2 متصل می شود. غشای ER، این منجر به تغییر در ساختار پروتئین و کانال باز می شود - CaI+ وارد سیتوزول می شود. در غیاب IP3 در سیتوزول، کانال بسته می شود فعال شدن پروتئین کیناز C. * افزایش غلظت CaI+ در سیتوزول سلول، سرعت برهمکنش CaI+ با آنزیم سیتوزولی غیر فعال پروتئین کیناز C را افزایش می دهد. PKC) و پروتئین کالمودولین، بنابراین سیگنال دریافتی توسط گیرنده سلولی منشعب می شود. * اتصال پروتئین کیناز C با یون های کلسیم به آنزیم اجازه می دهد تا با مولکول های فسفولیپید غشای اسیدی، فسفاتیدیل سرین (PS) وارد برهمکنش با واسطه کلسیم شود. ). دی آسیل گلیسرول، که مراکز خاصی را در پروتئین کیناز C اشغال می کند، میل ترکیبی آن را برای یون های کلسیم بیشتر می کند. یک کمپلکس آنزیمی در سمت داخلی غشاء تشکیل می شود - [PKS] [CaI+] [DAG] [PS] - پروتئین کیناز C فعال، آنزیم های خاص فسفریله کننده در سرین و ترئونین.
دخالت پروتئین کالمودولین در انتقال سیگنال اینوزیتول فسفات
در سلول های بسیاری از بافت ها پروتئینی به نام کالمودولین وجود دارد که به عنوان گیرنده CaI+ داخل سلولی عمل می کند؛ این پروتئین دارای 4 مرکز برای اتصال CaI+ است. کمپلکس [کالمودولین] - فعالیت آنزیمی ندارد، اما برهمکنش کمپلکس با پروتئین ها و آنزیم های مختلف منجر به فعال شدن آنها می شود.
خود تنظیمی سیستم
مانند بسیاری از سیستم های سیگنال دهی غشایی، سیستم فسفات اینوزیتول نه تنها یک مکانیسم تقویت، بلکه یک مکانیسم سرکوب سیگنال نیز دارد. اینوزیتول 1،4،5-تری فسفات (IP3) موجود در سیتوزول و دی آسیل گلیسرول (DAG) در غشاء می توانند در نتیجه یک سری واکنش ها دوباره به فسفاتیدیل 4،5-بیس فسفات (PIF2) تبدیل شوند. کاتالیز کاهش فسفولیپید توسط پروتئین فسفوریلاسیون کیناز C فعال می شود. غلظت CaI+ در سلول تحت تأثیر CaI+ ATPases غشای سیتوپلاسمی و ER و همچنین ترانسلوکازهای Na+/CaI+ و H+/CaI+ به سطح اولیه کاهش می یابد (فعال). آنتی پورت) غشای سلولی و میتوکندری. ترانسلوکازهای فعال CaI+ و CaI+-ATPases را می توان با: کمپلکس [کامودولین]، پروتئین کیناز A (فسفوریلاسیون)، پروتئین کیناز C (فسفوریلاسیون) فعال کرد. کاهش غلظت Ca2 در سلول و دی اسیل گلیسرول در غشاء منجر به تغییر در ترکیب پروتئین کیناز C، کاهش میل ترکیبی آن برای فسفاتیدیل سرین می شود، آنزیم به سیتوزول (شکل غیر فعال) تجزیه می شود. آنزیم ها و پروتئین های فسفریله شده توسط پروتئین کیناز C به دفسفریله تبدیل می شوند تحت تأثیر فسفوپروتئین فسفاتاز تشکیل می شود.
b-subunit: خصوصیات عمومی
زیرواحد b نقش عمده ای در عملکرد پروتئین های G ایفا می کند. GTP را متصل می کند. این به طور برگشت پذیر با زیر واحدهای c و d تعامل دارد، زمانی که تولید ناخالص داخلی در مرکز است به آنها می پیوندد و زمانی که GTP در مرکز قرار دارد از هم جدا می شود. هنگامی که GTP b متصل می شود، زیر واحد فعال می شود و توانایی تنظیم سیستم های موثر درون سلولی را به دست می آورد. زیر واحدهای b برخی از پروتئینهای G میتوانند دستخوش تغییرات شیمیایی شوند. تحت تأثیر سموم وبا و سیاه سرفه، پروتئین ها با پی دی اف ریبولیزاسیون در باقی مانده های آرژنین و سیستئین در انتهای C رخ می دهد که در نتیجه عملکرد طبیعی پروتئین های G مختل می شود. علاوه بر این، پروتئین کیناز C می تواند زیرواحد b پروتئین G خالص شده و پروتئین Gz در داخل بدن را فسفریله کند. ظاهراً پروتئین ها در تمام این موارد غیر فعال هستند.بیشتر پروتئین های G دارای زیرواحدهای b با وزن مولکولی حدود 40 کیلو دالتون هستند.
زیر واحدهای c و d: خصوصیات کلی
زیرواحدهای بتا و گاما یک کمپلکس با یکدیگر تشکیل می دهند که تنها در شرایط دناتوره شدن از هم می پاشند. نقش آنها کاملاً مشخص نیست. در آزمایشات با ترانسدوسین و سپس با پروتئین Gi، نشان داده شد که زیر واحدهای بتا و گاما برای برهمکنش پروتئین G با گیرنده و جایگزینی GDP با GTP ضروری هستند. کمپلکس بتا گاما به شدت با غشاء و به عنوان لنگر برای زیرواحد b عمل می کند. هنگامی که زیرواحد ب بتا گاما جدا می شود، کمپلکس بتا گاما می تواند به سیتوپلاسم منتقل شود.علاوه بر اتصال و مهار فعالیت زیرواحد ب بتا گاما، در برخی موارد اثر مستقیم بر روی فاکتور دارد. سیستم های سلولی فسفولیپاز A2 را فعال می کند و با کالمودولین تعامل می کند و در نتیجه فعالیت آدنیلات سیکلاز مغز را مهار می کند. کمپلکس G-بتا-گاما تحریک AC1 توسط Gs-alpha را مهار می کند.AC2 با اتصال G-بتا-گاما تحریک می شود، اما فقط در حضور Gs-alpha.AC3 نیز توسط G-بتا-گاما تحریک می شود. کانال های پتاسیم قلب با اتصال Gs-alpha و G-beta-gama باز می شوند. غشای سلولی: تحریک هیدرولیز فسفولیپید. غشای سلولی: تغییر در محتوای cAMP.
G-proteins: g-sub واحدها
فسفوریلاسیون گیرنده ها یکی از مکانیسم های تنظیم فعالیت آنهاست. زیرواحدهای rg پروتئین های G می توانند با فعال کردن پروتئین کینازهایی که گیرنده ها را فسفریله می کنند، بازخورد منفی ایجاد کنند. این پروتئین کینازها GRK نامیده می شوند. پروتئین کینازهای GRK شامل رودوپسین کیناز و بتا آدرنرژیک کیناز است. فسفوریلاسیون منجر به حذف گیرنده کیناز می شود. به عنوان مثال، گیرنده های موسکارینی و آدرنرژیک فسفریله شده در سرین و ترئونین در دامنه C ترمینال با اتصال آریستین هدف قرار می گیرند، که آنها را برای حذف توسط اندوسیتوز آغاز می کند. به طور معمول، C-پایانه گیرنده دارای چندین مکان برای فسفوریلاسیون توسط پروتئین کینازهای مختلف است. مشخص شده است که یک محرک ضعیف (غلظت کم آگونیست) پروتئین کیناز A را فعال می کند و یک محرک قوی پروتئین کیناز b-ARK را فعال می کند که با فسفریله کردن گیرنده، انتقال سیگنال به آدنیلات سیکلاز را قطع کرده و تولید cAMP را متوقف می کند. فسفوریلاسیون توسط پروتئین کیناز A زمانی اتفاق می افتد که 10% از گیرنده ها اشغال شده باشند. در این حالت، فسفوریلاسیون سایر گیرندههای اشغال نشده منجر به آزادسازی زیرواحدهای b و فسفوریلاسیون مربوطه توسط پروتئین کیناز دیگری b-ARK میشود. زیرواحدهای b-ARK محلیسازی، اتصال مؤثر و غیرفعال کردن زیرواحدهای b را تضمین میکنند و میل گیرندهها را تنظیم میکنند. برای لیگاندهای فعال کننده آنها و کاهش توانایی GDR برای تفکیک از زیر واحد (تثبیت حالت غیرفعال)، کانال K + موسکارینی را در قلب باز می کند، کانال Ca2+ را در غشای پیش سیناپسی می بندد، فسفولیپاز PLA2 و برخی ایزوفرم های فسفولیپاز را فعال می کند. C، میل ترکیبی گیرنده را برای آگونیست تنظیم می کند GTP (GTP) - پروتئین های اتصال دهنده انتقال سیگنال از گیرنده ها به سیستم های موثر درون سلولی با استفاده از پروتئین های متصل شونده به GTP انجام می شود. پروتئین های متصل شونده به GTP با استفاده از مکانیسم خاصی - چرخه فاز GTP - فعال/غیرفعال می شوند.
پروتئین های متصل شونده به GTP دو خانواده اصلی از پروتئین های G و پروتئین های با وزن مولکولی کم GTP را تشکیل می دهند.
همه این پروتئین ها به شدت GTP را متصل می کنند و آن را به GDP تبدیل می کنند و در این حالت پروتئین از حالت فعال به حالت غیرفعال تبدیل می شود. خواص پروتئین های متصل شونده به GTP ویژگی ساختاری اصلی پروتئین های متصل شونده به GTP وجود یک دامنه اتصال نوکلئوتید گوانین است. وجود توالیهای آمینو اسیدی مورد توافق، نشانهای واضح است که پروتئین متعلق به این خانواده است.
ادبیات
1. بیوشیمی. ویرایش سوم (تجدیدنظر شده). M; گروه انتشارات GEOTAR-Media. 20062. مبانی بیوشیمی. A. White, F Handler, E. Smith, R. Hill, I. Lehman.3 جلد M; جهان 19813. بیوشیمی. موری، گرنر. M; مدرسه تحصیلات تکمیلی. 19934. بیوشیمی. لنینگر، 19955. بیوشیمی. V.P. کوموف.، V.N. شودووا. بوستارد; م: 2004 .6. بیوشیمی عمومی کتاب درسی بیوشیمی. دوره سخنرانی. M.T. جنگین; 19977. مبانی بیوشیمی. یو.بی. فیلیپوویچ. چاپ دوم، بازنگری و گسترش یافته است. مدرسه تحصیلات تکمیلی؛ M4، 1995
(گیرنده های جفت شده با پروتئین C، GPCRs)، سیگنالی را از پیام رسان های اولیه به اهداف درون سلولی با استفاده از آبشار پروتئین GPCR-^-G-protein^-effector منتقل می کنند. سیگنال های اولیه برای این گیرنده ها طیف گسترده ای از مولکول ها، به عنوان مثال، هورمون ها و انتقال دهنده های عصبی با وزن مولکولی کم (مانند آدرنالین، نوراپی نفرین، استیل کولین، سروتونین، هیستامین)، مواد افیونی، پپتید و هورمون های پروتئینی (آدرنوکورتیکوتروپین، سوماواستاتین، آنژیو پرسین، وازوپرسین). گنادوتروپین، فاکتور رشد اپیدرمی)، برخی نوروپپتیدها.
همین سری شامل سیگنال های شیمیایی بسیاری است که توسط سلول های حسی بویایی و چشایی درک می شوند و نور، گیرنده آن رنگدانه سلول های بینایی یا گیرنده نوری رودوپسین است.
باید در نظر گرفت که یک سیگنال اولیه می تواند انتقال سیگنال را از طریق چندین (گاهی بیش از 10) GPCR مختلف آغاز کند، بنابراین اگر تعداد سیگنال های خارجی برای GPCR ها چندین ده باشد، خود بیش از 200 گیرنده از این قبیل شناخته شده اند.
GPCR ها با همه تنوعشان، پروتئین های غشایی انتگرال مونومری هستند که زنجیره پلی پپتیدی آن هفت بار از غشای سلولی عبور می کند. در همه موارد، محل گیرنده مسئول برهمکنش با سیگنال اولیه در سمت بیرونی غشاء و ناحیه در تماس با پروتئین G در سمت سیتوپلاسمی آن قرار دارد.
جزء پایین دست آبشار انتقال سیگنال GPCR توسط پروتئین G نشان داده می شود. حدود 20 پروتئين G مختلف يافت شده است كه در ميان آنها ابتدا بايد به Gs و C اشاره كرد كه به ترتيب آدنيلات سيكلاز را تحريك و مهار مي كنند. Gq، فعال کننده فسفولیپاز C. پروتئین های G سلول های حسی: گیرنده نوری - G t (ترانسدوسین)، بویایی - G o if و چشایی - G g.
پروتئینهای C هتروتیمرهایی هستند که از سه نوع زیر واحد تشکیل شدهاند: ct، (S و y)، اما در شرایط طبیعی دو زیرواحد آخر بهعنوان یک کمپلکس منفرد Ru عمل میکنند. مهمترین ویژگی پروتئینهای C وجود یک مرکز اتصال guani بر روی زیرواحد a آنها - نوکلئوتیدها: GDP و GTP (شکل های 139، 145). سپس این شکل مربوط به حالت غیر فعال پروتئین است (شکل 79).
رویداد اصلی در انتقال یک سیگنال از گیرنده، که تحت تاثیر سیگنال اولیه، به پروتئین G است، این است که گیرنده فعال، تبادل GDP متصل به پروتئین G را برای GTP موجود در محیط کاتالیز می کند. این تبادل GDP/GTP روی پروتئین G با تفکیک مولکول پروتئین G trimeric به دو زیر واحد عملکردی همراه است: زیرواحد α حاوی GTP و کمپلکس Py (شکل های 139، 145).
در مرحله بعد، یکی از این زیرواحدهای عملکردی، که به نوع سیستم سیگنال دهی بستگی دارد، با پروتئین مؤثری که توسط یک آنزیم یا کانال یونی نمایش داده می شود، تعامل دارد. در نتیجه، فعالیت کاتالیزوری یا هدایت یونی آنها بر این اساس تغییر می کند، که به نوبه خود منجر به تغییر در غلظت سیتوپلاسمی پیام رسان ثانویه (یا یون) می شود و در نهایت، یک یا آن پاسخ سلولی را آغاز می کند.
پروتئین های موثر در سیستم های سیگنال دهی مانند پروتئین GPCR-G-protein-effector می توانند آدنیلات سیکلاز باشند که سنتز cAMP از ATP را کاتالیز می کند. فسفولیپاز C، که فسفاتیدیلینوزیتول را هیدرولیز می کند تا DAT و 1P3 را تشکیل دهد. فسفودی استراز، که cGMP را به GMP تجزیه می کند. برخی از انواع کانال های پتاسیم و کلسیم.
مهم است که در طول انتقال سیگنال در آبشار گیرنده پروتئین G-protein-effector، سیگنال خارجی اصلی را می توان چندین برابر تقویت (تقویت) کرد. این به دلیل این واقعیت است که یک مولکول گیرنده در حالی که در حالت فعال (R*) است، موفق می شود چندین مولکول پروتئین G را به شکل فعال شده (G*) تبدیل کند.
به عنوان مثال، در آبشار بصری، رودوپسین^C^ecGMP-فسفودی استراز برای هر مولکول R* می تواند چندین صد یا حتی هزاران مولکول Gt* تولید کند، به این معنی که در مرحله اول آبشار 7?*-»G* ضریب تقویت سیگنال خارجی 10 2 -10 3 است. اگرچه در مرحله بعدی آبشار (پروتئین موثر C*^) هر مولکول G* تنها با یک مولکول از پروتئین مؤثر برهمکنش میکند، سیگنال در اینجا نیز تقویت میشود، زیرا برای هر مولکول G* و بر این اساس، از ماده فعال شده پروتئین عامل، بسیاری از آنها در مولکول های پیام رسان ثانویه سیتوپلاسم ظاهر می شوند (یا ناپدید می شوند). بنابراین، در آبشار بصری در مرحله دوم، یک مولکول cGMP فسفودی استراز فعال شده قادر است تا 3000 مولکول cGMP را در هر ثانیه تجزیه کند، که به عنوان یک پیام رسان ثانویه در سلول های گیرنده نوری عمل می کند.
سود کل آبشار برابر است با حاصلضرب بهره در تمام مراحل آبشار. ضریب تقویت سیگنال هنگام عبور از آبشار می تواند به مقادیر بسیار بالایی برسد: در سلول های بصری این مقدار از مرتبه 10 5 -10 6 است.
توقف محرک خارجی با خاموش شدن تمام اجزای سیستم سیگنالینگ همراه است. در سطح گیرنده، اولاً در نتیجه تفکیک پیام رسان اولیه از کمپلکس با GPCR و ثانیاً با فسفوریلاسیون گیرنده ها تحت تأثیر پروتئین کینازهای خاص و متعاقباً اتصال به گیرنده اصلاح شده یک خاص به دست می آید. پروتئین (به عنوان مثال، P-arrestin).
پروتئین های G دارای فعالیت GTPase هستند، یعنی توانایی هیدرولیز GTP مرتبط با آنها به تولید ناخالص داخلی، که حذف خود را تضمین می کند، یعنی انتقال G-GTP-e G-GDP. از آنجایی که وضعیت فعال شدن پروتئین افکتور (روشن-خاموش) مستقیماً به وضعیت پروتئین G بستگی دارد، این انتقال همچنین به معنای خاموش شدن پروتئین مؤثر و در نتیجه توقف سنتز (هیدرولیز) پروتئین است. پیام رسان ثانویه یا بسته شدن کانال یونی.
و در نهایت، برای اینکه انتقال سلول به حالت اولیه (قبل از محرک خارجی) کامل شود، مکانیسمهای خاصی سطح اولیه پیامرسان یا کاتیون ثانویه را در سیتوپلاسم آن باز میگردانند. به عنوان مثال، cAMP، که غلظت سیتوپلاسمی آن در طول انتقال سیگنال در آبشار گیرنده P-آدرنرژیک افزایش مییابد C5-protein-adenylate cyclase، سپس توسط cAMP فسفودی استراز به AMP غیر حلقوی (خطی) هیدرولیز میشود، که این ویژگی را ندارد. یک پیام رسان ثانویه
در میان مواد آلی سنجاب ها، یا پروتئین هابیوپلیمرهای متعدد، متنوع ترین و دارای اهمیت بسیار بالایی هستند. سهم آنهاست 50 - 80%
توده سلولی خشک
مولکول های پروتئین از نظر اندازه بزرگ هستند، به همین دلیل به آنها می گویند درشت مولکول ها. علاوه بر کربن، اکسیژن، هیدروژن و نیتروژن، پروتئین ها ممکن است حاوی گوگرد، فسفر و آهن باشند. پروتئین ها از نظر تعداد (از صد تا چند هزار)، ترکیب و توالی مونومرها با یکدیگر تفاوت دارند. مونومرهای پروتئینی اسیدهای آمینه هستند (شکل 1)
تنوع بی نهایتی از پروتئین ها با ترکیب های مختلف همه چیز ایجاد می شود 20
آمینو اسید. هر اسید آمینه نام، ساختار و خواص خاص خود را دارد. فرمول کلی آنها را می توان به صورت زیر ارائه کرد:
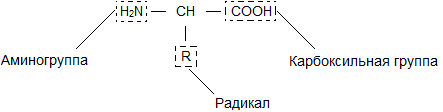
یک مولکول آمینو اسید از دو بخش یکسان با تمام اسیدهای آمینه تشکیل شده است که یکی از آنها گروه آمینه است. -NH 2) با خواص پایه، دیگری با یک گروه کربوکسیل ( -کوه) با خاصیت اسیدی. بخشی از یک مولکول به نام رادیکال ( آر) اسیدهای آمینه مختلف ساختارهای متفاوتی دارند. وجود گروه های بازی و اسیدی در یک مولکول اسید آمینه، واکنش پذیری بالای آنها را تعیین می کند. اسیدهای آمینه از طریق این گروه ها ترکیب می شوند و پروتئین ها را تشکیل می دهند. در این حالت، یک مولکول آب ظاهر می شود و الکترون های آزاد شده یک پیوند پپتیدی تشکیل می دهند. به همین دلیل پروتئین نامیده می شود پلی پپتیدها.
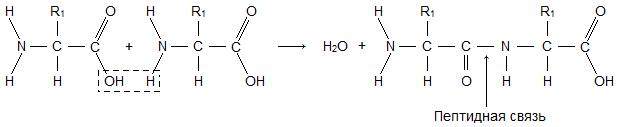
مولکول های پروتئین می توانند پیکربندی های فضایی متفاوتی داشته باشند و در ساختار آنها چهار سطح سازماندهی ساختاری وجود دارد.
توالی اسیدهای آمینه در یک زنجیره پلی پپتیدی است ساختار اولیهسنجاب این برای هر پروتئینی منحصر به فرد است و شکل، خواص و عملکرد آن را تعیین می کند.
اکثر پروتئین ها در نتیجه تشکیل پیوندهای هیدروژنی بین آنها شکل مارپیچی دارند -CO-و -NH-گروه هایی از باقی مانده های اسید آمینه مختلف زنجیره پلی پپتیدی. پیوندهای هیدروژنی ضعیف هستند، اما در مجموع ساختار نسبتاً محکمی ایجاد می کنند. این مارپیچ است ساختار ثانویهسنجاب
ساختار سوم- "بسته بندی" فضایی سه بعدی زنجیره پلی پپتیدی. نتیجه یک پیکربندی عجیب اما خاص برای هر پروتئین است - گلبول. استحکام ساختار سوم توسط پیوندهای مختلفی که بین رادیکال های اسید آمینه ایجاد می شود تضمین می شود.
ساختار کواترنریبرای همه پروتئین ها معمول نیست. در نتیجه ترکیب چندین ماکرومولکول با ساختار سوم در یک مجتمع پیچیده ایجاد می شود. به عنوان مثال، هموگلوبین در خون انسان مجموعه ای از چهار ماکرومولکول پروتئینی است.
این پیچیدگی ساختار مولکول های پروتئین با تنوع عملکردهای ذاتی این پلیمرهای زیستی مرتبط است.
نقض ساختار طبیعی یک پروتئین نامیده می شود دناتوره سازی. این می تواند تحت تأثیر دما، مواد شیمیایی، انرژی تابشی و عوامل دیگر رخ دهد. با یک ضربه ضعیف، فقط ساختار چهارتایی متلاشی می شود، با ساختار قوی تر - سوم، و سپس ثانویه، و پروتئین به شکل یک زنجیره پلی پپتیدی باقی می ماند.
این فرآیند تا حدی قابل برگشت است: اگر ساختار اولیه آسیب نبیند، پروتئین دناتوره شده قادر به بازیابی ساختار خود است. نتیجه این است که تمام ویژگی های ساختاری یک ماکرومولکول پروتئین توسط ساختار اولیه آن تعیین می شود.
بجز پروتئین های ساده، که فقط از اسیدهای آمینه تشکیل شده است، نیز وجود دارد پروتئین های پیچیده
به روز رسانی: 1393/01/15
103583
اگر متوجه خطایی شدید، یک متن را انتخاب کنید و Ctrl+Enter را فشار دهید







